I could of course harp on now about how important photosynthesis is to all life on Earth and how we couldn't live without it. Of course, this is true. But you probably know that. Oh, and don't forget this: water + carbon dioxide ----> glucose + oxygen. Let's dive straight in to the chloroplast.
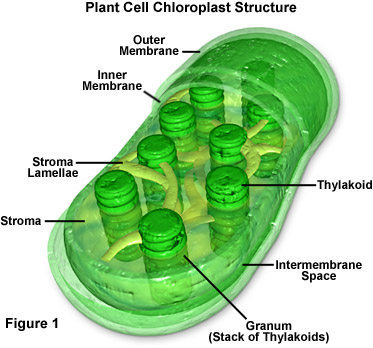
Photosynthesis occurs in two stages: the light dependent reaction (LDR) comes first and occurs on the thylakoid membrane of the chloroplasts, then comes the light independent reaction (LIR) which can also be called the Calvin cycle which occurs in the stroma of the chloroplast. It is also important to note the main structures in the chloroplast and their function:
- The chloroplast envelope. A double-membrane enclosure which runs around the whole chloroplast. The two membranes create a intermembrane space between them.
- The thylakoids are small disks which contain photosystems 1 and 2 (PSI and PSII) on their membrane to capture light energy and utilise it in the LDR. A stack of thylakoids is called a granum (pl. grana).
- Each granum becomes a part of an interconnected network with lamellae connecting grana between thylakoids.
- The stroma is a complex 'soup' of enzymes (for the LIR), starch grains (stores of glucose), sugars, salts and organic acids.
In my (slightly dry) blog on ATP, I mentioned that photosynthesis is a process which both uses and produces ATP. This results in a net production of ATP of 0. It is the LDR which produces this ATP for its brother in arms, the LIR, to use in synthesising glucose. In some ways, the LDR can be thought of as the LIR's helper in that it produces two important products for it. These are ATP and reduced NADP (a photosynthetic coenzyme). The LDR is made up of three main steps:
- Photolysis (catalytic breakdown using light) of water
- Reduced NADP production
- ATP production

The image above shows a model of the LDR and I suggest (strongly, very strongly because I will go off on tangents about nothing to do with this) that you use it to see how far along in the whole process we are. The sequence occurs from left (PSII) to right (ATP synthase).
We'll start at the beginning at step 1. PSII contains an enzyme which can split water into its three constituent parts: half an oxygen molecule (one O atom), two protons and two electrons. These protons and electrons will be used later, but the oxygen is released as a waste product. This is known as photolysis. However, the enzyme needs help from light to do this (hence: photo-lysis). If we can synthesis similar enzymes or inorganic catalysts, we could use them to split water into hydrogen and oxygen for use in fuel cells to make cars which only emit water as a product of use (how very considerate).
Now to step 2 (which is quite a bit longer). Two photons (particles of light energy) hit PSII which contains an array of pigments which can trap the energy of these photons. These pigments come under two categories:
- accessory pigments (xanthophyll, carotene...), which transfer the light energy to the primary pigment.
- the primary pigment (chlorophyll), which uses the energy to promote a pair of electrons (not in a you're getting a big office way - more physics than business)
These two electrons are promoted to a higher energy level (which means the electrons have more energy or are 'excited'). In their excited state, they have enough energy to escape the primary pigment. They then pass through an electron transport chain (ETC) consisting of proton pumps which use the energy of the electrons to pump protons (from photolysis) into the thylakoid lumen - creating an 'electrochemical gradient' between the thylakoid lumen and the stroma. This becomes useful later. Finally, after all that proton pumping, the electrons reach a final electron acceptor at the end of the ETC. But if all those electrons keep leaving the primary pigment, won't it run out? Nope. Remember the electrons released from water in photolysis? These are used to replenish the electrons lost from PSII or to 'reduce' the pigment. Remember that OILRIG (oxidation is loss, reduction is gain).
Once the electrons have reached the end of the ETC and the final electron acceptor, they reduce PSI. Why does it need reducing? Because two more photons excite two more electrons from PSI. These electrons are used to achieve the final goal of step two of the LDR - to reduce NADP to become reduced NADP. Each electron from the pair of promoted electrons is used, with a proton, to form reduced NADP.
NADP + proton + electron ----> reduced NADP
So, moving on to step 3. The protons pumped into the thylakoid in step 2 are now used. Because of all the positive charge and concentration of protons which has built up, the protons want to diffuse out of the thylakoid. The only way they can do this, is through the enzyme ATP synthase. In diffusing out of the thylakoid, they must donate their energy to ATP synthase which uses this energy to form ATP from ADP and an inorganic phosphate group. The movement of these protons is called chemiosmosis.
The whole process above is called non-cyclic photophosporylation. There is another version of the LDR called cyclic photophosphorylation where PSII isn't used, so the electrons promoted from PSI come back to reduce PSI back to its original state. This doesn't involve photolysis of water or reduced NADP production. It only produces small amounts of ATP, so is less preferable compared with non-cyclic photophosphorylation.
There we have the LDR, in all three stages, producing lots of ATP and reduced NADP for the LIR, which we are about to investigate.
The LIR, carbon fixation, or Calvin cycle (discovered by a scientist called Melvin Calvin - the best scientist name ever) is the next process in photosynthesis. It uses the products of the LDR with carbon dioxide to produce glucose, or in fact a whole range of organic compounds, which the plant can use later on. Below is a diagram of the Calvin cycle.

Here's a description of what's happening (the names I use don't always match up with the ones on the diagram):
- Carbon dioxide which enters the leaf through the stomata reacts with ribulose bisphosphate (RuBP) in the presence of the enzyme, rubisco to form 2 molecules of glycerate 3-phosphate (GP).
- 2 ATP molecules and 2 reduced NADP molecules react with the 2 GP molecules to form 2 triose phosphate (TP) molecules.
- 5 in 6 molecules of TP then react with an ATP molecule to reform RuBP and the cycle restarts.
- The other molecule of TP then goes to form useful organic products. If we use glucose as an example, two TP molecules are needed to make a molecule of glucose (because TP has 3 carbons and glucose has 6).
All of the reacted ATP and reduced NADP then return to the LDR to be reformed back to ATP and reduced NADP.
Because only 1/6 of all TP produced is used to make organic products, the Calvin cycle must occur 6 times to produce one molecule of glucose. This is because each carbon dioxide which is input into the cycle results in the production of 2 TP molecules. If 1/6 are formed into useful products, we need 12 TPs to make one glucose: 12 x 1/6 = 2.
Even though photosynthesis is a brilliant process which fuels all life on Earth, it can be limited by environmental conditions. The most important ones are:
- Light intensity
- Light wavelength
- Carbon dioxide concentration
- Water availability
- Temperature
If we take a look at light intensity, having a light intensity too low would result in not enough photons reaching the thylakoid membrane for a high rate of reaction for the LDR. If not enough reduced NADP and ATP can be produced, then this will also limit the rate of the LIR.
Wavelength is also a very important consideration of the light used for photosynthesis. Each pigment will only use light of a certain wavelength. This is why plants contain several different pigments; so more wavelengths of light can be used for the LDR. Despite this, not all wavelengths are suitable.
Obviously, carbon dioxide concentration is very important in photosynthesis. If it is too low, the Calvin cycle cannot occur quickly and this limits the overall rate of photosynthesis. However, an excess of carbon dioxide can cause poisoning to the plant, so the carbon dioxide concentration must be high, but not too high.
Water availability is less of an issue for plants, so long as there is some. If not enough water is available for non-cyclic photophosphorylation to occur, cyclic photophosphorylation can be used. A complete absence of water, would however result in the death of the plant.
As with all enzyme-catalysed reactions, if the temperature is too low, the rate of reaction will be low. If the temperature is too high, the enzyme will be denatured and the reaction cannot be catalysed. This is all about active sites, frequency of collision and particle energy.
Even if you were to have optimum conditions for all of these factors, photosynthesis cannot occur at an infinite rate, clearly. So there will always be one factor which limits the others. No matter how much you increase the other factors, the limiting factor will dictate the rate of photosynthesis.
And there we have photosynthesis. Even though it is a lot more complex than you might think, it does mean that we and all other life can survive, so we'll forgive it just this once.